
Why do trees take more risks in the Amazon? <sup>†</sup>
Abstract
Rowland et al. (2015) published recently in Nature a study suggesting that "death from drought in tropical forests is triggered by hydraulics not carbon starvation". We have reanalyzed they data with the objective of placing them in a more global perspective.
Why do trees take more risks in the Amazon?
Drought is one of the major forces shaping forest ecosystems around the world, but much remains unknown about the mechanisms by which drought causes trees to die (McDowell et al. 2011, Sevanto et al. 2014, Sala et al 2010, Salmon et al. 2015). Rowland et al. (2015) have significantly advanced our understanding in this area. They showed that hydraulic failure of the xylem tissue, rather than carbon starvation, was the main factor triggering tree death in the tropical rainforest. The tree species of this ecosystem generally operate beyond the limits of hydraulic failure, which leaves them highly vulnerable to drought-induced death. This discovery is particularly significant, because hydraulic failure is highly predictable as it is determined solely by the structure of the sap transport system of each species and drought intensity.
Here, we aim to place the key findings of Rowland et al. (2015) into a more global perspective. The hydraulic safety margin (HSM) of trees can be defined as the difference between the minimum xylem pressure experienced during drought and the pressure causing 50% hydraulic failure. For this study, we computed the HSM values from a xylem functional trait database (Choat et al. 2012) covering 681 plant species from different sites and forest biomes worldwide. The HSM data for the Brazilian Amazon were obtained from Rowland et al. (2015).
The HSM values reported for the different forest biomes studied worldwide to date have always been positive, suggesting that hydraulic failure occurs only under conditions of extreme water deficit (Delzon and Cochard 2014). The HSM values of the species studied by Rowland et al. (2015) are unique in being very narrow in a control plot and even largely negative in a through-fall exclusion experiment (Fig. 1). This indicates that these trees operate at xylem pressures very close to the point of no return. The vulnerability of their xylem tissue is remarkably high, much higher than reported for all the other biomes of the planet Choat et al. 2012). It will be important to confirm these values as the air-injection technique used in this study is known to overestimate xylem vulnerability for species with long vessels (Cochard et al. 2013, Torres-Ruiz et al. 2014). Similarly, the relationship found by Rowland et al. (2015) between xylem vulnerability and tree size might be biased by an open vessel artifact as taller trees tend to have longer vessels.
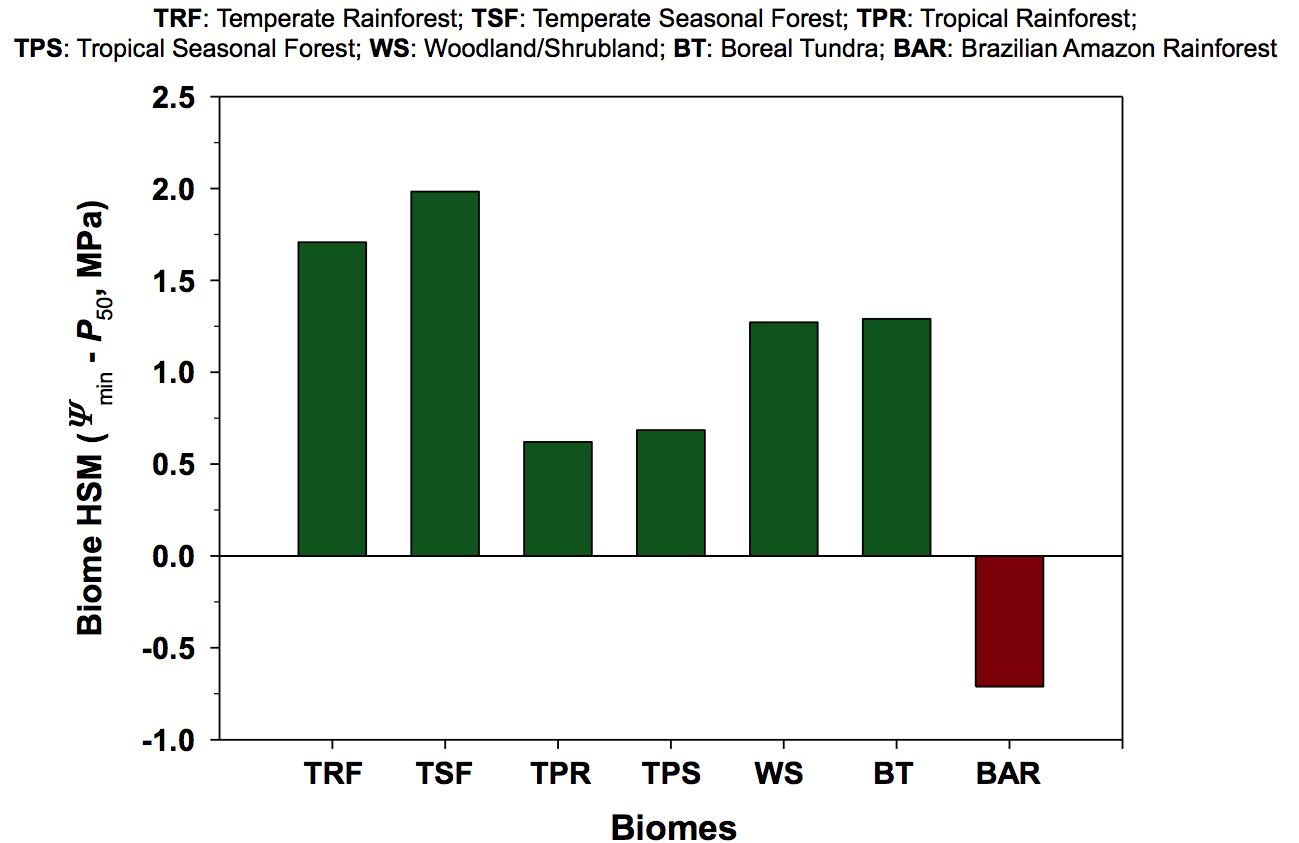
Figure 1. Hydraulic safety margins (HSM) for different forest biomes (green columns from Choat et al., 2012) and for the species studied by Rowland et al. (2015) in the tropical rainforest of the Brazilian Amazon (dark red column).
If correct, this study raises many fascinating questions. Why do trees in the tropical rainforest of Brazilian Amazonia live so close to the edge? How do they cope with such small safety margins and high levels of embolism on a daily and seasonal basis? Will tree mortality rates increase in Amazonia with the predicted increase in drought event frequency due to climate change (IPCC 2007)? Further studies will be required to answer these questions, but a key point will be determining whether the findings of Rowland et al. (2015) apply to the entire Amazonian Basin. This is particularly relevant given that significant decreases in the size of the tropical rainforest in Amazonia, the largest in the world, would lead to major changes not only in the carbon cycle (Pan et al. 2011), but also in global atmospheric circulation, which is powered by the evaporation and condensation processes occurring within the rainforest (Malhi et al. 2007).
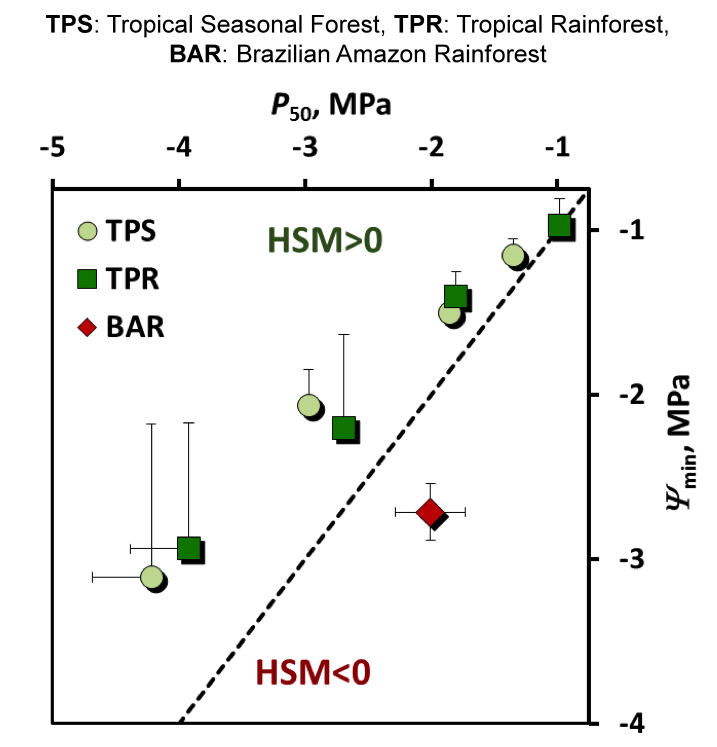
Figure 2. HSM values for different tropical biomes worldwide (dark and light green symbols) and for the Brazilian Amazon (dark red diamond). Ψmin: minimum xylem pressure experienced during drought; P50: pressure causing 50% hydraulic failure. Species in the Brazilian rainforest operate with negative safety margins, leaving them highly vulnerable to drought-induced death.
Very few HSM values have been reported for tropical species, but the available information (Choat et al. 2012, Barigah et al. 2014) suggests that other tropical biomes operate with safer margins (Fig. 2). The rather complex history of the Amazonian forest during recent glacial periods has shaped a mosaic of different habitats and ecosystems (Leite et al. 2016). The patterns observed by Rowland et al. (2015) in the Caxiuanã National Forest Reserve may not, therefore, be representative of the whole basin. A more global survey of tree hydraulic traits for the entire Amazon forest will be required to predict the stability of this ecosystem in the near future. Fortunately, robust, practical techniques are now available for large-scale surveys of this type Cochard et al. 2005, 2015).
† Unrefereed comment
References
- Barigah TS, Aussenac G, Baraloto C, Bonal D, Cochard H, Granier A, Guehl JM, Huc R, Sobrado MA, Tyree MT. 2014. The water relations of two tropical rainforest species (Virola surinamensis and Eperua falcata): Is Virola unusual as previously reported? Journal of Plant Hydraulics 1: e002.
- Choat B, Jansen S, Brodribb TJ, Cochard H, Delzon S et al. 2012. Global convergence in the vulnerability of forests to drought. Nature 491: 752–55. doi:10.1038/nature11688
- Cochard, Damour G, Bodet C, Tharwat I, Poirier M, Améglio T. 2005. Evaluation of a new centrifuge technique for rapid generation of xylem vulnerability curves. Physiol Plantarum 124: 410–418. doi:10.1111/j.1399-3054.2005.00526.x
- Cochard H, Badel E, Herbette S, Delzon S, Choat B, Jansen S. 2013. Methods for measuring plant vulnerability to cavitation: a critical review. Journal of Experimental Botany 64: 4779-479. doi:10.1093/jxb/ert193
- Cochard H, Delzon S, Badel E. 2015. X-ray microtomography (micro-CT): a reference technology for high-resolution quantification of xylem embolism in trees. Plant Cell and Environment. 38: 201–206. doi:10.1111/pce.12391
- Delzon S, Cochard H. 2014. Recent advances in tree hydraulics highlight the ecological significance of the hydraulic safety margin. New Phytologist 203: 355–358. doi:10.1111/nph.12798
- IPCC Climate Change. 2007. The Physical Science Basis. Vol 1009. Cambridge University Press, Cambridge, UK.
- Leite YL, Costa LP, Loss AC, Rocha RG, Batalha-Filho H, Bastos AC, et al. 2016. Neotropical forest expansion during the last glacial period challenges refuge hypothesis. Proceedings of the National Academy of Sciences 201513062. doi:10.1073/pnas.1513062113
- Malhi Y, Roberts JT, Betts RA, Killeen TJ, Li W, Nobre CA. 2008. Climate change, deforestation, and the fate of the Amazon. Science 319: 169–172. doi:10.1126/science.1146961
- McDowell NG. 2011. Mechanisms linking drought, hydraulics, carbon metabolism, and vegetation mortality. Plant Physiology 155: 1051–1059. doi:10.1104/pp.110.170704
- Pan Y, Birdsey RA, Fang J, Houghton R, Kauppi PE, Kurz WA et al. 2011. A large and persistent carbon sink in the world’s forests. Science, 333: 988–993. doi:10.1126/science.1201609
- Rowland L, Da Costa ACL, Galbraith DR, Oliveira RS, Binks OJ, Oliveira AAR et al. 2015. Death from drought in tropical forests is triggered by hydraulics not carbon starvation. Nature 528:119–122. doi:10.1038/nature15539
- Sala A, Piper F, Hoch G. 2010. Physiological mechanisms of drought-induced tree mortality are far from being resolved. New Phytologist 186: 274–281. doi:10.1111/j.1469-8137.2009.03167.x
- Salmon Y, Torres-Ruiz JM, Poyatos R, Martinez-Vilalta J, Meir P, Cochard H, Mencuccini M. 2015. Balancing the risks of hydraulic failure and carbon starvation: a twig scale analysis in declining Scots pine. Plant Cell and Environment 38: 2575–2588. doi:10.1111/pce.12572
- Sevanto S, McDowell NG, Dickman LT, Pangle R, Pockman WT. 2014. How do trees die? A test of the hydraulic failure and carbon starvation hypotheses. Plant Cell and Environment 37: 153–151. doi:10.1111/pce.12141
- Torres-Ruiz JM, Cochard H, Mayr S, Beikircher B, Diaz-Espejo A, Rodriguez-Dominguez CM, Badel E, Fernández JE. 2014. Vulnerability to cavitation in Olea europaea current-year shoots: further evidence of an open-vessel artifact associated with centrifuge and air-injection techniques. Physiologia Plantarum 152: 465–474. doi:10.1111/ppl.12185
Attachments
No supporting information for this articleArticle statistics
 Views: 2842
Views: 2842
Downloads
 PDF: 561
PDF: 561