
Evidence for Air-Seeding: Watching the Formation of Embolism in Conifer Xylem
Abstract
Water transport in plants is based on a metastable system as the xylem “works” at negative water potentials (ψ). At critically low ψ, water columns can break and cause embolism. According to the air-seeding hypothesis, this occurs by air entry via the pits. We studied the formation of embolism in dehydrating xylem sections of Juniperus virginiana (Cupressaceae), which were monitored microscopically and via ultrasonic emission analyses. After replacement of water by air in outer tracheid layers, a complex movement of air-water menisci into tracheids was found. With decreasing ψ, pits started to aspirate and the speed of menisci movements increased. In one experiment, an air-seeding event could be detected at a pit. The onset of ultrasonic activity was observed when pits started to close, and ultrasonic emission ceased at intense dehydration. Experiments clearly indicated that predictions of the air-seeding hypothesis are correct: At low ψ, pit mechanisms to prevent air entry failed and air spread into tracheids. ψ fluctuations caused complex movements of air-water menisci and pits, and at low ψ, air-seeding caused ultrasonic emissions. Main insights are presented in a video.
Introduction
Water transport in plants is based on a cheap but risky mechanism. Negative pressure (i.e. low water potential ψ) caused by the transpiration is transmitted via continuous water columns to the roots and the soil (cohesion-tension theory; Boehm 1893, Dixon and Joly 1894). The water columns thereby are in a metastable state as low ψ can induce a phase transition from fluid water to vapour and, in consequence, cause a breakage of water columns (i.e. embolism). The ψ thresholds for embolism formation are species-specific with overall higher resistances in drought adapted plants (e.g. Brodribb 2009, Choat et al. 2012).
The formation of embolism is probably not induced by a homogenous nucleation within the water column but by a heterogenous nucleation according to the “air-seeding hypothesis” (Crombie et al. 1985, Sperry and Tyree 1988, Cochard et al. 1992, Tyree et al. 1994, Tyree and Zimmemann 2002): At critically low ψ, air is pulled into water-filled conduits from adjacent, air-filled areas, such as intercellular spaces, mechanically damaged conduits or embolised conduits. In consequence, air can spread from conduit to conduit. This probably occurs via the pits, which connect the conduits. In angiosperms, the pit membrane is porous and the dimension of pores in the pit membrane defines the critical threshold for air-seeding (Tyree et al. 1994). Small pores stabilize the air-water interface at higher ψ gradients so that the largest pore of a conduit determines its air-seeding resistance. Conifers evolved special valve-like pit structures. The pit membrane is composed of a central thickening (the torus) surrounded by a net with wide pores (the margo). When ψ gradients rise, the torus is aspirated to the pore of the pit chamber, which isolates adjacent tracheids and prevents air entry. At critical ψ, this mechanism fails leading to air-seeding and formation of embolism (see e.g. Hacke and Sperry 2001, Tyree and Zimmemann 2002, Pittermann et al. 2005). Critical ψ thresholds thus depend on the pit architecture (e.g. Cochard et al. 2009, Pittermann et al. 2010, Delzon et al. 2010, Jansen et al. 2012, Bouche et al. 2014).
There are numerous indications that embolism formation in fact is based on air-seeding (e.g. Sperry and Tyree 1990, Cochard et al. 1992) but, to our knowledge, no direct observation of air-seeding was published up to now. This is probably due to the fact that any manipulation of the xylem system can disturb the metastable state of water columns and cause artificial embolism so that insights into xylem are difficult. In the present study, we tried to enable these insights by use of thin xylem sections and microscopic observations as in experiments presented by Lewis et al. (1994) and combined these observations with a monitoring of acoustic emissions (e.g. Tyree and Dixon 1983, Kikuta 2003, Mayr and Rosner 2011). Also Kikuta et al. (2003) used conifer sections, which were dehydrated over solutions of low water potential and monitored via an ultrasonic sensor and a dew point hygrometer. We followed the dehydration of conifer xylem on air and produced a video (see supplement airseeding.mpg), which shows the process of air entry into tracheids.
Materials and Methods
Experiments were performed on stem sections of Juniperus virginiana L. growing in the garden of the Institute of Botany, University of Natural Resources and Life Sciences, BOKU Vienna. This species lacks resin channels in the wood and therefore is best suitable for observations of the tracheid system.
Radial cross sections (ca. 25 x 5 mm) with a thickness of 100 µm were prepared with a microtome (Schlittenmikrotom OME, Reichert, Wien, Austria). It can be estimated that each cross section contained about ten layers of tracheids. While most of the outermost tracheids were probably opened by cutting, the central tracheid layers were expected to be intact. The sections were thin enough to allow focusing on the inner layers and watching air-water interfaces in the respective tracheids. Sections were stored in distilled water until use in experiments.
Sections were placed on an object slide and kept wet until the following installations were finished:
(i) One side (ca. 10 mm) of the section was covered by an ultrasonic sensor (R15/C 150 kHz resonance sensor, 80-400 kHz) connected to a 20/40/60 dB preamplifier (set to 40 dB) and an ultrasonic emission analysis system (PCI-2 ultrasonic system PAC 125 18-bit A/D, 3 kHz-3 MHz PCI2; all components: Physical Acoustics, Wolfegg, Germany). The threshold was set to 28 dB, peak definition time, hit definition time and hit lockout time were 200 µs, 400 µs and 2 µs, respectively. Registration and analysis of ultrasonic acoustic events (UAE) were done with AEwin software (Physical Acoustics). Signals were transformed to audible frequencies (audio monitor option) and recorded in an audio file during the experiment.
(ii) The object slide was then transferred to a microscope (Olympus BX 41, system microscope, Olympus Austria, Vienna, Austria) and the part of the section, which was not covered by the ultrasonic sensor, placed under the objective at 4x, 10x or 20x magnification. The focus was set to tracheids in the centre of the section before a video camera (Sony DCR-HC39E, Sony, Vienna, Austria) was positioned at the ocular via a stative. Furthermore, the microscope was interfaced with a digital camera (Sony Cyber-shot, DSC-W17, Vienna, Austria) and a monitor to control picture quality and focus during the experiment.
The section was then allowed to dehydrate in air. Please note that no cover slip was used to enable desiccation. Unavoidably, this caused the quality of pictures to decrease during the experiment. The experiment was finished when the section was completely dehydrated. The experiment was repeated several times and representative video sequences were used to produce the film (airseeding.mpg) available in the supplement. In addition, two videoframe sequences were extracted and one record of ultrasonic emissions is given with the text.
Results
The following description of results refers to the film available in the supplement (airseeding.mpg): After a short introduction demonstrating the experimental setup, the main sequence of this video (timeframe in video 0´52-5´32) shows a dehydrating section with the respective acoustic emissions. Inserted film sequences show details with higher magnification, whereby tracheids are always oriented horizontally. The last insert shows the waveforms of ultrasonic emissions (4´42).
Main sequence (0´52-5´32)
At the beginning of this sequence (0´52-1´20), air entered the space below the section. This occurred rapidly in relatively big areas and caused slight shifts in the color. Afterwards, air started to enter tracheids, whereby menisci of the air-water interface moved slowly at the beginning but speeded up with ongoing dehydration. The movement of air into tracheids occurred in both directions, and sometimes simultaneous movements in opposite directions were observed. First acoustic signals were registered at 3´29 and the maximum of acoustic emissions around 4´40 (Fig. 1). At this stage, movement of air through tracheids occurred at high speeds so that the shift of menisci could hardly be followed. The resolution of pictures decreased significantly after about 5´, when most of the section was air-filled. The video ends after the last acoustic events detected. Small movements of the section and the autofocus function of the camera caused some small artificial zoom movements within the film sequence.
Inserts
Sequence 1 (1´38): air enters tracheids
This sequence shows the start of air entry into the tracheids, first probably into conduits cut open by the preparation. Air-water menisci showed no continuous movement through tracheids but a stepwise shift, interrupted by stops and even short drawbacks (also see Fig. 2). As observed in the main sequence, the movement direction differed between tracheids.
Sequence 2 (2´10) and sequence 3 (2´40, high resolution): pit aspiration
When the section was sufficiently dehydrated (probably when all tracheids cut open were air-filled), ψ gradients between air-filled and water-filled tracheids built up and caused aspiration of pits. Pit closure can be seen by movements and contrast changes in the pit chambers, which are due to air entry at one side of the pit. According to the complex movements of air menisci described above, pits did not close at once but showed a fluctuation between aspiration and opening until they closed permanently.
Sequence 4 (3´50): air-seeding
In one experiment, we were lucky to focus on a pit, at which air-seeding occurred (Fig. 3). A series of similar pits showed pit torus movements and aspiration as in the previous sequences. Then, an air bubble suddenly appeared at the pit pore and enlarged around the pit. After a period without changes (cut in video) further enlargement and air movement through the tracheid occurred stepwise and in both directions.
Discussion
The experiments enabled the visualization of several processes driven by dehydration. During the first minutes of dehydration, it was eye-catching that the air entered tracheids stepwise with many forward and backward movements (Fig. 2). This was due to ψ changes in the tracheid network. When air entered a tracheid, the tension in this and adjacent conduits was released and ψ increased until dehydration induced the next drop in ψ. This capacitive effect was already described by several authors (e.g. Tyree and Yang 1990, LoGullo and Salleo 1992, Höltta et al. 2009). It should also be noted that the direction of air movements was not homogenous. It depended on the point of air entry, if the meniscus moved in basal or distal direction. Embolism thus may not only expand in axial direction but also spread transverse to the grain, as indicated by embolism patterns observed in some cross sections (e.g. Tyree and Zimmermann 2002, Mayr et al. 2006). The time required for dehydration and the patterns of embolised conduits within samples were in accordance with the study of Lewis et al. (1994), who monitored embolism formation in Thuja occidentalis sections. The video also shows the speed of menisci movements to increase with ongoing dehydration. We suggest that the initial spreading of air was rather artificial and caused by air entry into tracheid with ends cut open by the preparation. Later, intact tracheid layers in the sample center were affected and this process probably occurred similar to the process in natura.
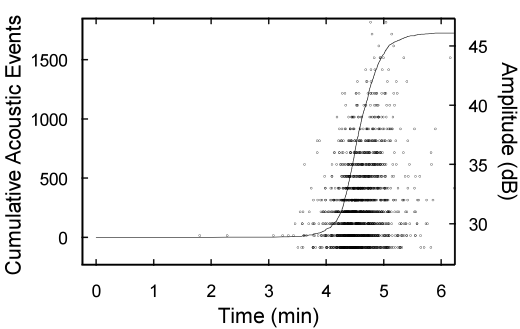
Figure 1: Ultrasonic emissions during dehydration. Cumulative acoustic events (solid line) as well as the amplitude of signals (circles) are given.
The pits of these intact tracheids obviously aspirated (e.g. Pittermann et al. 2005) and withstood ψ differences. Similar to complex movements of menisci in the tracheids, ψ fluctuations in the xylem caused the tori to move several times until they closed permanently. Then, half of the pit chamber was air-filled, while the other half and the respective tracheid was still water-filled. Recently, it was demonstrated that the ratio of aspirated pits correlates well with the ratio of embolised tracheids in Picea abies xylem affected by winter-drought and freeze-thaw induced embolism (Mayr et al. 2014).
According to the air-seeding hypothesis, air should spread across pits when a critical ψ gradient is reached. We were able to observe this process only one time (Fig. 3); as each tracheid contains numerous pits, one has to be very lucky focusing on the pit which shows air-seeding. Nevertheless, our observation clearly indicates that the air-seeding hypothesis correctly describes the induction of embolism. Unfortunately, the time resolution of our video did not allow unraveling the exact entry point of air. Although it is still unclear if air-seeding across conifer pits is related to torus displacement, seal capillary seeding or torus capillary seeding (Tyree et al. 1994; Hacke and Sperry 2001; Domec et al. 2006; Cochard et al. 2009; Hacke and Jansen 2009; Delzon et al. 2010; Jansen et al. 2012, Plavcova et al. 2014) the video is a strong evidence that embolism formation is based on air-seeding.
The main sequence in the video also shows that ultrasonic emissions started relatively late during dehydration, although air movement was observed from the beginning on. It is likely that air-seeding occurred before acoustic signals could be detected. As the energy of ultrasonic signals depends on the size of embolising conduits and the tensile forces acting on the cell walls (Mayr and Rosner 2011), ψ in the system was probably too moderate to produce detectable signals during initial dehydration stages.

Figure 2: Air movement in tracheids. Pictures (a-j) were extracted from a video sequence in 1s intervals. Air-water menisci in tracheids at the upper right corner (tracheid rows above arrow) show forward and backward movements.
In contrast, numerous signals of increasing intensity (Fig. 1; after ca. 4 min.) were detected, when pit aspiration, air-seeding and a fast movement of many air-water menisci was observed. This pattern also corresponds to the three phases of air-seeding postulated by Fanyi et al. (2012) with acoustic emissions only on rapid expansion at low ψ. On further dehydration, ultrasonic activity ceased because all tracheids were embolised. This shows that ultrasonic emission analysis allow to follow embolism formation “on-line” but it cannot be expected that each air-seeding event produces one signal.
Kikuta et al. (2003) demonstrated that xylem sections embolised at less negative ψ than intact branches. It thus should be considered that the size of xylem portions and of air-water interfaces substantially influences the dynamics and risk of embolism formation (Kikuta et al. 2003, Mayr et al. 2006). Nevertheless, we believe that wood sections are a useful tool to study air-seeding at the level of tracheids and pits and that observed processes correspond to air-seeding in living plants. Our observations clearly indicated that predictions of the air-seeding hypothesis are correct. It is possible to explain observed hydraulic processes based on the structural and functional aspects of this important hypothesis. It might be promising to repeat the experiments with a high-speed camera and study the dynamics and mechanisms of air-seeding in more detail.
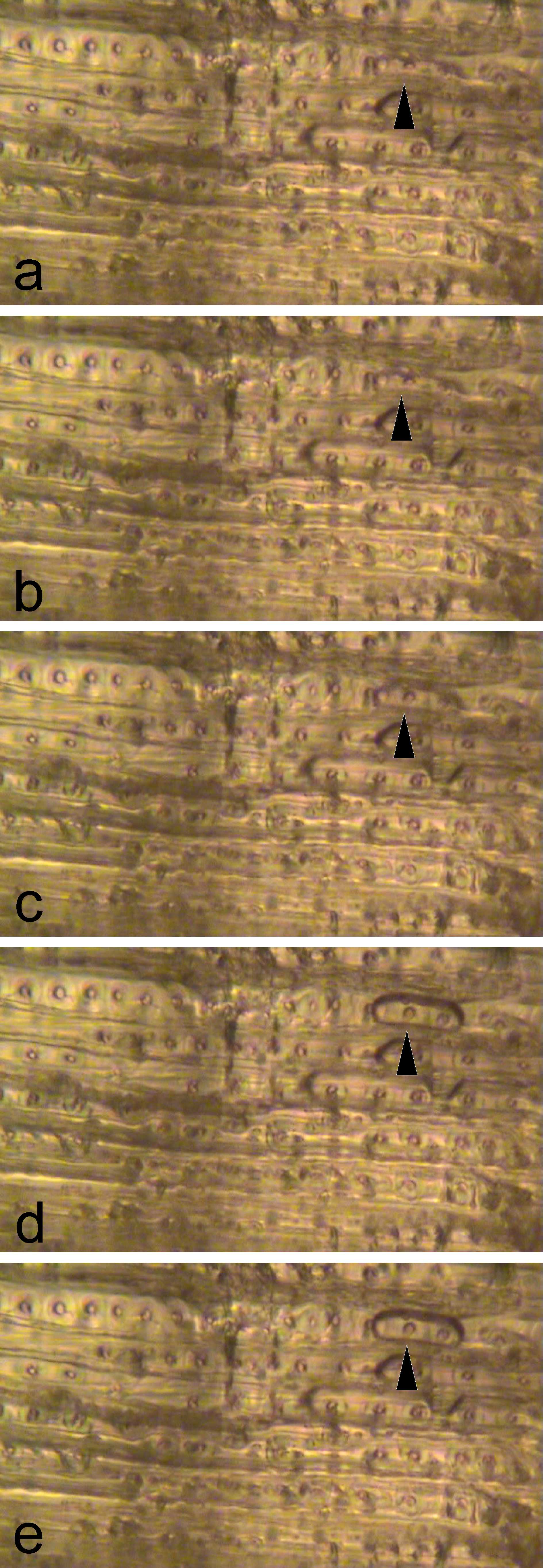
Figure 3: Air-seeding. Pictures (a-e) show consecutive frames (interval 0.04 s) extracted from a video sequence. At one pit at the upper right corner (arrow), an air bubble enters the respective tracheid.
Acknowledgements : we thank Mag. Birgit Dämon for helpful assistance and the ÖKOM company, Austria for providing the camera equipment. This study was supported by the Austrian Science Fund (FWF), project I826-B25.
References
- Boehm H. 1893. Capillarität und Saftsteigen. Berichte der Deutschen Botanischen Gesellschaft 11: 203-212.
- Bouche PS, Larter M, Domec J-C, Burlett R, Gasson P, Jansen J, Delzon S. 2014. A broad survey of hydraulic and mechanical safety in the xylem of conifers. Journal of Experimental Botany 65: 4419-4431 doi:10.1093/jxb/eru218
- Brodribb TJ. 2009. Xylem hydraulic physiology: The functional backbone of terrestrial plant productivity. Plant Science 177: 245-251 doi:10.1016/j.plantsci.2009.06.001
- Choat B, Jansen S, Brodribb TJ, Cochard H, Delzon S, Bhaskar R, Bucci SJ, Field TS, Gleason SM, Hacke UG, Jacobsen AL, Lens F, Maherali H, Martínez-Vilalta J, Mayr S, Mencuccini M, Mitchell PJ, Nardini A, Pittermann J, Pratt RB, Sperry JS, Westoby M, Wright IJ, Zanne AE. 2012. Global convergence in the vulnerability of forests to drought. Nature 491: 752-755 doi:10.1038/nature11688
- Cochard H, Cruiziat P, Tyree MT. 1992. Use of positive pressure to establish vulnerability curves: further support for the air-seeding hypothesis and implications for pressure-volume analyses. Plant Physiology 100: 205-209 doi:10.1104/pp.100.1.205
- Cochard H, Hölttä T, Herbette S, Delzon S, Mencuccini M. 2009. New insights into the mechanisms of water-stress-induced cavitation in conifers. Plant Physiology 151: 949–954 doi:10.1104/pp.109.138305
- Crombie DS, Hipkins MF, Milburn JA. 1985. Gas penetration of pit membranes in the xylem of Rhododendron as the cause of acoustically detectable sap cavitation. Australian Journal of Plant Physiology 12: 445-453 doi:10.1071/PP9850445
- Delzon S, Douthe C, Sala A, Cochard H. 2010. Mechanism of water-stress induced cavitation in conifers: bordered pit structure and function support the hypothesis of seal capillary-seeding. Plant, Cell and Environment 33: 2101-2111 doi:10.1111/j.1365-3040.2010.02208.x
- Dixon HH, Joly J. 1894. On the ascent of sap. Proceedings of the Royal Society London 57: 3-5 doi:10.1098/rspl.1894.0126
- Domec JC, Lachenbruch B, Meinzer FC. 2006. Bordered pit structure and function determine spatial patterns of air-seeding thresholds in xylem of Douglas-Fir (Pseudotsuga menziesii; Pinaceae) trees. American Journal of Botany 93: 1588-1600 doi:10.3732/ajb.93.11.1588
- Fanyi S, Yuansheng W, Yanxia C, Li Z. 2012. Three types of cavitation caused by air seeding. Tree Physiology 32: 1413-1419 doi:10.1093/treephys/tps089
- Hacke UG, Jansen S. 2009. Embolism resistance of three boreal conifer species varies with pit structure. New Phytologist 182: 675-686 doi:10.1111/j.1469-8137.2009.02783.x
- Hacke UG, Sperry JS. 2001. Functional and ecological xylem anatomy. Perspectives in Plant Ecology, Evolution and Systematics 4: 97-115 doi:10.1078/1433-8319-00017
- Hölttä T, Cochard H, Nikinmaa E, Mencuccini M. 2009. Capacitive effect of cavitation in xylem conduits: Results from a dynamic model. Plant, Cell and Environment 32: 10-21 doi:10.1111/j.1365-3040.2008.01894.x
- Jansen S, Lamy J-B, Burlett R, Cochard H, Gasson P, Delzon S. 2012. Plasmodesmatal pores in the torus of bordered pit membranes affect cavitation resistance of conifer xylem. Plant, Cell and Environment 35: 1109-1120 doi:10.1111/j.1365-3040.2011.02476.x
- Kikuta SB. 2003. Ultrasound acoustic emissions from bark samples differing in anatomical characteristics. Phyton 43: 161-178.
- Kikuta SB, Hietz P, Richter H. 2003. Vulnerability curves from conifer sapwood sections exposed over solutions with known water potential. Journal of Experimental Botany 54: 2149-2155 doi:10.1093/jxb/erg216
- Lewis AM, Harnden VD, Tyree MT. 1994. Collapse of water-stress emboli in the tracheids of Thuja occidentalis L. Plant Physiology 106: 1639-1646.
- LoGullo MA, Salleo. 1992. Water storage in the wood and xylem cavitation in 1-year-old twigs of Populus deltoides Bartr. Plant, Cell and Environment 15: 431–438 doi:10.1111/j.1365-3040.1992.tb00993.x
- Mayr S, Rosner S. 2011. Cavitation in dehydrating xylem of Picea abies: energy properties of ultrasonic emissions reflect tracheid dimensions. Tree Physiology 31: 59-67 doi:10.1093/treephys/tpq099
- Mayr S, Rothart B, Wolfschwenger M. 2006. Temporal and spatial pattern of embolism induced by pressure collar techniques in twigs of Picea abies. Journal of Experimental Botany 57: 3157-3163 doi:10.1093/jxb/erl077
- Mayr S, Schmid P, Laur J, Rosner S, Charra-Vaskou K, Dämon B, Hacke UG. 2014. Uptake of water via branches helps timberline conifers refill embolized xylem in late winter. Plant Physiology 164: 1731-1740 doi:10.1104/pp.114.236646
- Pittermann J, Choat B, Jansen S, Stuart S, Lynn L, Dawson TE. 2010. The relationships between xylem safety and hydraulic efficiency in the Cupressaceae: the evolution of pit membrane form and function. Plant Physiology 153: 1919–1931 doi:10.1104/pp.110.158824
- Pittermann J, Sperry JS, Hacke UG, Wheeler JK, Sikkema EH. 2005. Torus-margo pits help conifers compete with angiosperms. Science 310: 1924-1924 doi:10.1126/science.1120479
- Plavcová L, Jansen S, Klepsch M, Hacke UG. 2014. Nobody's perfect: Can irregularities in pit structure influence vulnerability to cavitation? Frontiers in Plant Science. doi:10.3389/fpls.2013.00453
- Sperry JS, Tyree MT. 1988. Mechanism of water stress-induced xylem embolism. Plant Physiology 88: 581-587 doi:10.1104/pp.88.3.581
- Sperry JS, Tyree MT. 1990. Water-stress-induced xylem embolism in three species of conifers. Plant, Cell and Environment 13: 427-436 doi:10.1111/j.1365-3040.1990.tb01319.x
- Tyree MT, Dixon MA. 1983. Cavitation events in Thuja occidentalis L.? Ultrasonic acoustic emissions from the sapwood can be measured. Plant Physiology 72: 1094-1099 doi:10.1104/pp.72.4.1094
- Tyree MT, Davis SD, Cochard H. 1994. Biophysical perspectives of xylem evolution: Is there a tradeoff of hydraulic efficiency for vulnerability to dysfunction? IAWA Journal 15: 335-360 doi:10.1163/22941932-90001369
- Tyree MT, Yang S. 1990. Water-storage capacity of Thuja, Tsuga and Acer stems measured by dehydration isotherms. Planta 182: 420–426 doi:10.1007/BF02411394
- Tyree MT, Zimmermann MH. 2002. Xylem structure and the ascent of sap. 2nd edition, Springer, Berlin, pp 331 doi:10.1007/978-3-662-04931-0
Attachments
Research Instrument
Mayr S, Kartusch B, Kikuta S. 2014.
Evidence for air-seeding: watching the formation of embolism in conifer xylem.
Article statistics
 Views: 5484
Views: 5484
Downloads
 PDF: 465
PDF: 465